水产品在储藏过程中保持其风味品质对其市场价值和消费者接受度至关重要。本研究评价了壳聚糖-表没食子儿茶素没食子酸酯涂层对鲟鱼鱼片风味的影响,并探讨了其潜在的调节机制。感官评价、电子鼻和挥发性化合物和三磷酸腺苷相关化合物的分析表明,涂层显著减少了异味成分,改善了风味质量。涂层抑制了游离氨基酸和游离脂肪酸水平,抑制了风味转换酶的活性,减缓了脂肪代谢。相关分析突出了关键风味化合物与风味前体、酶和代谢物之间的正相关关系。高通量测序确定乳球菌、沙雷氏菌和气单胞菌是影响风味特征的关键微生物。这些发现促进了对风味恶化机理的理解,并提出了在冷藏过程中保持鲟鱼鱼片风味质量的潜在策略。
鲟鱼(Aristichthys Nobilis)以其优质的肉类而闻名,其特点是营养丰富,味道细腻。然而,鲜鱼比许多其他食物腐烂得更快,这对保持市场价值构成了挑战。鲤鱼风味品质下降的主要原因是微生物的生长和酶活性的提高。假单胞菌、沙雷氏菌和气单胞菌等微生物代谢鱼中的营养物质,产生异味并影响鱼的风味质量。同时,与风味有关的酶活性,如涉及脂肪酶、蛋白酶和核苷酸酶的酶活性,会加速脂肪、蛋白质和与ATP相关的化合物的分解,导致不愉快的风味物质的积累。蛋白质代谢和脂肪降解产生游离氨基酸(FAA)和游离脂肪酸(FFA),进而产生挥发性风味化合物。具体地说,脂肪氧合酶(LOX)将游离脂肪酸转化为醛,然后乙醇脱氢酶(ADH)将醛转化为醇。最终,酒精酰基转移酶(AAT)利用这些醇来生成酯。然而,一些高级醇、醛、羧酸和酯是从FAA的代谢中衍生出来的。此外,碱性磷酸酶(AKP)、酸性磷酸酶(ACP)和5‘-核苷酸酶(5’-NT)催化umamitast IMP的分解,导致苦味和不利的Flavors HxR和Hx的产生。因此,研究如何通过控制腐败微生物和降低酶活性来延缓鲤鱼鱼肉风味的恶化,对水产工业的发展具有潜在的价值。表没食子儿茶素没食子酸酯(EGCG)是一种有效的儿茶素,主要存在于绿茶中,以其强大的抗菌和抗氧化特性而闻名。EGCG通过破坏细菌细胞膜和抑制基本细菌酶而显示出广谱的抗菌活性。EGCG主要通过与活性部位结合、形成稳定的EGCG-酶复合体和螯合必需的金属离子来抑制酶的活性。这些作用分别阻止底物进入、诱导构象改变和剥夺酶的必要辅助因子,从而降低或停止酶的功能。因此,EGCG具有巨大的潜力。水产品保鲜。近年来,使用EGCG增强的可食用涂层显示出优异的抗菌和抗氧化性能,有效地延长了水产品的货架期。同时,含有EGCG的涂层消除了在食品中直接添加游离的EGCG,同时提供了受控释放,因为它逐渐从薄膜中释放EGCG。因此,选择一种可食用、可持续、经济的涂膜基质材料是扩大EGCG在水产品中应用的关键。壳聚糖是最丰富、最可再生的多糖之一,具有良好的成膜性、抗菌性和抗氧化性,非常适合作为保鲜涂料的基材。壳聚糖的抗菌活性源于它能够分解微生物细胞壁中的肽聚糖,导致细胞内电解质渗漏,最终导致细胞死亡。一些研究使用了含有生物活性化合物的壳聚糖基涂料来保存水产品,如金凤尾和草鱼,实现了有效的腐败抑制和货架期延长。然而,据我们所知,很少有研究探讨壳聚糖-EGCG涂层对冷却鲤鱼肉整体风味代谢的影响,特别是关于其与高通量测序、风味相关酶活性和代谢组学相关的潜在调控机制。我们推测壳聚糖-EGCG涂层可以减少冷藏鲤鱼鱼片中异味化合物的产生,从而改善其整体风味品质。此外,我们假设这种改善是通过抑制与风味相关的酶的活性,抑制腐败微生物的增殖,以及减缓蛋白质、脂肪和核苷酸的降解来实现的。因此,我们的工作旨在通过结合风味转换酶活性、高通量测序和代谢组学来评估壳聚糖-EGCG涂层对鲤鱼鱼片风味代谢的影响,并探讨其潜在的调控机制。通过电子鼻和感官评定分析其感官特性。详细监测包括挥发性风味化合物、FAA、FFA、ATP相关化合物和风味转换酶活性。高通量测序测量了微生物组成的变化,而代谢组学则分析了代谢途径的变化。通过皮尔逊和曼特尔的测试,阐明了风味变化与酶活性、腐败微生物和代谢物之间的关系。这项工作对风味恶化的机制提供了创新的见解,并提出了保持水产品风味质量的潜在调控策略。
样品处理:以醋酸和甘油(AG)为包衣液,将1g醋酸和1g甘油溶解于100mL超纯水中。将2g壳聚糖溶于100mL1%冰醋酸中,在25℃下搅拌2 h后,加入1g甘油作为增塑剂,再搅拌30min,制得壳聚糖(CH)包衣溶液。壳聚糖-表没食子儿茶素没食子酸酯(CE)包衣溶液的制备方法与此类似,每100毫升溶液中含有2克壳聚糖、1克醋酸和1克甘油。充分搅拌后,加入1.5g的EGCG,在黑暗中再搅拌2小时。涂布前,将所有包衣溶液冷却至4℃。将准备好的鱼片随机分配到四组,其中三组在4℃下浸泡在AG、CH和CE包衣溶液中,每组1分钟。然后在25℃的无菌通风区域烘干20分钟,以允许薄膜形成。未处理组作为对照组(CON)。所有样品都进行真空包装,在4℃下保存15天。到那时,没有涂层的鱼片已经开始显著变质,这是水产品研究中广泛报道的现象。每三天收集一次样本并进行分析,直到储存结束。每项测量至少进行一式三份。
重点内容
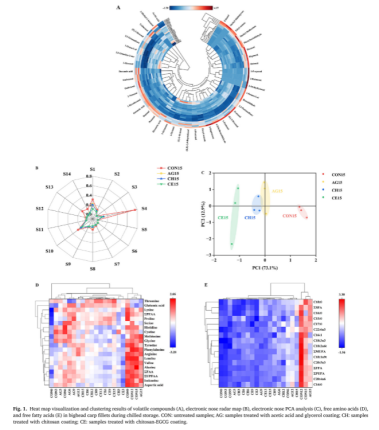
共鉴定出42种挥发性风味化合物,分类如下:醇(9)、酮(6)、醛(19)、酯(4)和有机酸(4)。热图被用来直观地展示挥发性风味化合物的变化(图1a)。贮藏15d后,CON组和AG组的总挥发性风味物质含量均明显增加,分别从474.13 ng/g上升到2511.78 ng/g和474.13 ng/g上升到1202.21 ng/g。然而,CH组和CE组的VFC浓度在整个储存过程中一直波动,分别为680.05 ng/g和374.84 ng/g。在贮藏过程中,CE组的挥发性风味化合物浓度始终最低,其次是CH、AG和CON组。这表明CE涂层提供了良好的风味保存,并有效地抑制了异味。醛是水产品中重要的挥发性风味化合物,因其阈值低、气味难闻而被认为是水产品中重要的挥发性风味物质。醛来源于不饱和脂肪酸的氧化、酶催化(特别是脂氧合酶,LOX)和微生物活动。壬醛以其强烈的油味和酸味而闻名,而辛醛则散发着青草的气味。庚醛和(E,E)-2,4-庚二烯醛散发出强烈的鱼腥味。此外,己醛是储存过程中样品中发现的主要醛,它既显示鱼腥味也显示草味。
电子鼻可以复制动物的嗅觉系统,通过专门的传感器和识别模块快速检测挥发性有机化合物。如图1B所示不同样品的传感器对紧张程度的响应差异很大,表明不同处理和贮藏阶段它们的风味特征有很大差异。特别是,CON组对传感器S1(氨、胺)、S2(硫化氢、硫化物)、S4(乙醇)、S8(氨、硫化氢、乙醇)、S12(一氧化碳、乙醇)和S13(一氧化碳、氢、氮氧化物)的响应强度最高,其次是AG、CH和CE组。这表明CE组产生的乙醇、含硫化合物、含氮化合物等的浓度最低,并且在整个储存过程中抑制异味形成的效果最好,这与GC-MS的结果一致。使用传感器读数进行主成分分析(PCA)。数据点之间的间隔表示样品之间的风味特征差异:较小的间隔表示较小的差异,而较大的间隔表示较大的差异。如图1C所示,来自不同处理的样品被很好地分开,突出了各种涂层对鱼片风味的显著影响,特别是CE涂层。这些结果可能是由于EGCG具有强大的抗氧化剂和抗菌活性,可以抑制脂肪氧化和腐败微生物的生长,从而最大限度地减少异味的形成。以往的研究将FAA的味道属性分为两类:令人愉快的游离氨基酸(PFAA),包括Thr、Glu、Asp、Ala、Gly、Ser、Pro和Arg;以及令人不快的游离氨基酸(UPFAA),包括Leu、Phe、Ile、Lys、Val、Met、Tyr、His和Cys。创建了一张热图来显示各种FAA的相对丰度(图1D)。在储存过程中,四组的PFAA浓度均有波动,但与最初的水平相比,所有组的浓度都显示出总体上升的趋势。值得注意的是,UPFAA浓度在所有组中都呈现持续上升的趋势,在贮藏结束时,CON、AG、CH和CE组的UPFAA浓度分别上升了53.81%、42.91%、29.62%和21.52%。AG组和CH组UPFAA的轻微下降可能是由于醋酸和壳聚糖的抗菌性能所致,它们部分地抑制了腐败微生物对蛋白质的降解。CE组的UPFAA浓度最低,说明EGCG显著提高了壳聚糖涂层的生物活性。EGCG中的生物活性羟基(-OH)和双键可破坏细胞膜,导致细胞内渗漏,最终导致细菌细胞死亡。游离脂肪酸(FFA)在脂肪氧合酶催化或自氧化作用下分解代谢成脂肪醇、醛、酮、酯等低阈值挥发性物质,产生鱼腥味和腐臭味。总体而言,所有组的总FFA水平呈现出参差不齐的变化模式,CE组在总FFA增加和减少方面的波动最小。在贮藏结束时,CON组、AG组、CH组和CE组的FFA浓度分别增加了1.96倍、1.92倍、1.59倍和1.32倍。用热图显示各种游离脂肪酸的相对浓度(图1E)。
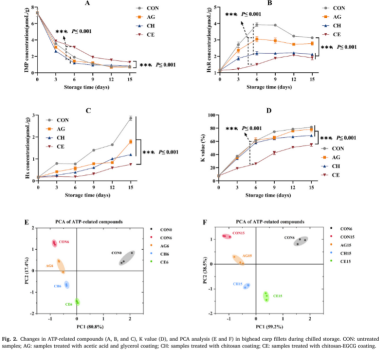
如图2a所示,IMP的初始浓度为7.31 Mol/g,与Yu等人的结果一致。所有组的IMP浓度显示出相似的模式:一开始迅速下降,然后逐渐下降,直到第15天。CE组在整个存储过程中保持了最高的IMP保留率。HxR浓度最初快速上升,随后略有下降,CE组在整个储藏过程中浓度最低。最初的上升可能是由于IMP的降解,而随后的下降是由于HxR水解为Hx。此外,在所有组中,HX浓度持续上升,其中CON组的浓度最高,其次是AG、CH和CE组。最近的报道表明,ATP转化为IMP可能完全通过自溶发生,而HxR和Hx的产生可能归因于腐败微生物的代谢活动和5’-NT、ACP和AKP等磷酸酶的催化。如图2E-F所示,主成分分析结果分别解释了总方差的98.20%和97.70%,表明样本在分布图上有明显的分离。一般来说,代表样本的点之间的距离越大,差异越明显,而距离越小,相似性越大。因此,CE组保存的新鲜度最接近原始样品,其次是CH、AG和CON组,进一步证明壳聚糖-EGCG涂层具有最好的保味和新鲜度。AG和CH组的轻微改善可能是由于醋酸和壳聚糖具有温和的酶抑制和抑菌活性。这些结果与先前关于冷藏期间含有葡萄籽提取物的罗非鱼片涂层明胶的报告一致。
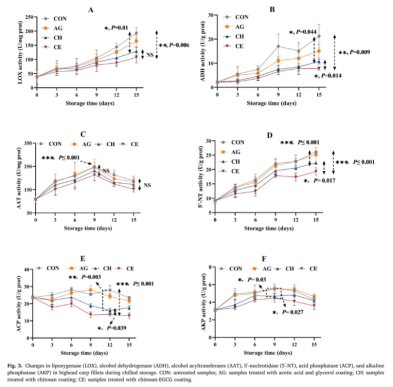
为了揭示壳聚糖-EGCG复合涂膜调味的内在机理,对几种关键酶的活性进行了分析。LOX是一种非血红素含铁脂肪酸双加氧酶,在水产品的酶促脂肪氧化中起关键作用。例如,亚麻酸在LOX的催化下,在C11和C12之间发生均质裂解,导致形成强烈鱼腥味的化合物(E,E)-2,4-七二烯。如图3A所示,在整个储存过程中,所有样品中的LOX活性都显著增加,在CON、AG、CH和CE组中分别增加了4.98、4.25、3.32和2.77倍。LOX活性的最初上升可归因于圆片引起的细胞破坏,导致LOX释放,而持续增加可能是由于LOX激活和特定腐败细菌的增殖。值得注意的是,CE组在整个储存过程中始终表现出最低的LOX活性,这可能是由于EGCG具有强大的抗菌和酶抑制特性。LOX催化的脂质氧化遵循自由基驱动的机制,酶活性部位的铁氧化脂肪酸,产生脂肪酸自由基和亚铁。然后,氧气与自由基中间体反应,形成一种脂肪酸氢过氧化氢,随后分解成带有鱼腥味的挥发性化合物。酚类化合物可以通过疏水相互作用或氢键结合到LOX活性部位,引起构象变化,从而降低LOX活性。此外,酚类化合物清除自由基的能力或它们与疏水脂肪酸底物的相互作用也会导致LOX活性降低。类似的结果也被报道,绿茶儿茶素对LOX催化的花生四烯酸和亚油酸的氧化有抑制作用。乙醇脱氢酶(ADH)在将醛转化为醇的过程中起着关键作用。图3B显示ADH活性的变化趋势与LOX相似,CON、AG、CH和CE组的ADH活性分别从2.18U/gprot增加到21.19U/gprot、15.06、10.73和7.79U/gprot。在贮藏过程中,CE组的ADH活性一直最低,其次是CH、AG和CON。这种模式与酒精挥发物,尤其是乙醇挥发物的水平和趋势相一致。以前的研究也表明ADH活性和酒精产量之间存在密切的相关性。乙醇酰基转移酶(AAT)是一种促进醇和酰基辅酶A结合,产生挥发性酯的酶,是一种重要的酶。在四个样本组中,AAT活性在最初的9天内稳步上升,然后在储存期的其余时间逐渐下降(图3C)。Zang等人在酸鱼发酵过程中观察到AAT活性变化的类似趋势。此外,在整个储藏期内,CE组的AAT活性最低,其次是CH、AG和CON,表明AAT活性的大小与酯水平之间存在明显的相关性。这些结果可能归因于EGCG对酶活性的强烈抑制作用。根据先前的研究,5‘-NT、ACP和AKP是水产品中IMP分解的主要酶。如图3D所示,5‘-NT活性在所有组均显著升高,至第15天,CON、AG、CH和CE的活性分别上升了2.86、2.76、2.44和2.11倍。然而,CON和AG组的ACP活性略有波动,但在贮藏期间保持相对稳定,最终分别达到23.45U/gprot和21.63U/gprot。总体而言,与初始值相比,CH组和CE组的ACP活性略有下降,最终分别降至17.66U/g prot和12.95U/g prot。此外,所有组的碱性磷酸酶浓度都表现出先略有上升后略有下降的趋势,在整个储藏期内活性变化很小。可以看出,在整个贮藏过程中,5‘-NT和ACP对IMP的降解起主导作用,而CE组的酶活性最低,这与观察到的IMP水平的变化趋势一致。鱼屠宰后,5‘-核苷酸酶、酸性磷酸酶、碱性磷酸酶活性的早期升高主要是由于溶酶体的分解,导致鱼组织中的酶释放。储藏期后期的上升与储藏期腐败微生物的增殖。因此,CE组的酶活性降低不仅归因于EGCG对内源酶的抑制,也归因于它对腐败微生物的抑制,这反过来又减少了细菌酶的产生。

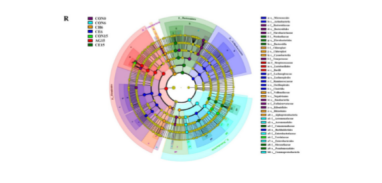

进行了微生物分析,以更好地了解CE涂层对微生物组成的影响。图4A显示,拟杆菌属、变形杆菌属、菲尔米特属和新球菌属是优势微生物种群,占总相对丰度的93%左右。变形杆菌和腐败菌已被确定为主要的腐败相关门,它们可能导致食品在储存过程中的风味恶化。随着贮藏时间的延长,CON、AG和CH组中变形杆菌和微生物的总比例呈上升趋势,从58.51%上升到99.00%、97.55%和76.62%。CE组变形杆菌和菌丝体的总比例略有上升,达到59.97%,最终下降到55.66%,说明EGCG的加入有效地提高了壳聚糖涂层的抗菌活性。在属水平上,最初的鱼片也表现出各种微生物特征,关键属包括链球菌属、小鼠李杆菌科、普雷沃特氏菌、不动杆菌属、真假单胞菌、类杆菌和假单胞菌(图4B)。随着贮藏时间的延长,四组鱼片中的细菌群落变得更加集中,鱼片中有少数几个属。储存6d后,乳球菌、假单胞菌、气单胞菌和沙雷氏菌成为CON组中的优势属,共占微生物群落总数的50.49%。值得注意的是,乳球菌的存在与脂肪氧合酶和酒精脱氢酶的活性密切相关,这对不同脂肪醇和醛的产生有直接影响。革兰氏阴性菌,包括气单胞菌和假单胞菌,与醇和醛的产生有关,特别是那些来自支链氨基酸的醇和醛的产生,例如2-甲基丁醇、3-甲基丁醇和3-甲基丁醛。沙雷氏菌是一种典型的腐败微生物,已知是某些醇(如1-辛烯-3-醇)和酯(如乙酸乙酯)的最高产生菌。在贮藏结束时,乳球菌、假单胞菌、气单胞菌和沙雷氏菌在CON、AG、CH和CE组中的组合比例分别达到72.96%、85.17%、31.47%和11.25%。含有EGCG的壳聚糖涂层显示出最强的抗菌活性,有效地抑制了产生异味挥发物的腐败微生物的繁殖,从而提高了冷藏过程中鱼片的风味品质。EGCG中的羟基促进了电子的离域,并作为质子供体,从而降低了细菌细胞膜的pH梯度。这种破坏削弱了质子的动力,耗尽了三磷酸腺苷,最终导致细胞死亡。此外,羟基可以通过与酶活性部位相互作用来改变微生物的代谢,从而抑制微生物的生长。进行了metaStat分析,基于前10个属的热图直观地显示了不同组之间的差异。如图4C所示,所有组的微生物多样性在储存结束时显著下降,乳球菌成为优势属。涂层鱼片中乳球菌的相对丰度显著下降,并有CH组与CE组之间差异有统计学意义。非度量多维标度(NMDS)被用来评估样本之间的差异,应力值低于0.2表示变化的准确表示(图4D)。随着距离的增加,各组样品与初始样品之间的微生物组成差异随着储存时间的延长而增大,其中CE组与原始样品的相似性最高。采用t检验评价微生物组成差异的显著性。如图4E-4H所示,乳球菌(p≤0.001)、沙雷氏菌(p≤0.01)和埃尔单胞菌(p≤0.05)的增长最为显著,而CE组这三个属受到显著抑制。Simpson和Shannon指数(图4I-4J)显示,贮藏后微生物多样性和均匀度普遍下降,而CE组在第15天达到最高水平,表明EGCG对维持物种多样性和均衡度具有积极作用。三元曲线图(图4K-4N)的结果表明,对照组乳球菌在贮藏后表现出最显著的增殖,而含有EGCG的壳聚糖涂层对乳球菌的抑制作用最强。此外,在第15天,与纯壳聚糖涂层相比,CE组对假单胞菌、链球菌和鼠李杆菌科的抑制效果更好。维恩图被用来说明常见和独特的扩增子序列变体(ASV)。如图4O和图P所示,在储存期内,常见ASV的数量从261个减少到50个,表明微生物群落结构发生了实质性变化。在贮藏结束时,对照组、CH组和CE组与新鲜样品之间的共同ASV分别为79、384和421,表明CE组的微生物群落结构与新鲜样品最相似。采用线性判别分析效应大小(LEFSE)分析,包括线性判别分析(LDA)评分和进化分支图,以确定具有统计学意义差异的生物标记。LDA得分在4以上的物种显示在LDA分布直方图中,其中条长表示它们不同影响的程度(图4Q)。进化分支图显示了从门到种的分类水平,由中心向外辐射的圆圈(图4R)。结果表明,在新鲜样品中,类杆菌和链球菌分别被鉴定为目和属水平的生物标志物。储藏15天后,沙雷氏菌和乳球菌。分别被认为是CON的属和种水平的生物标志物。对于AG,芽孢杆菌和乳球菌分别被鉴定为类级和属水平的生物标志物。对于CE,金黄色杆菌和不动杆菌是属水平的生物标志物,巴氏杆菌是类水平的生物标志物。因此,不同的贮藏时间和涂膜处理显著影响了鲤鱼的微生物群落组成和特征。添加了EGCG的壳聚糖包被显著抑制了主要腐败微生物乳球菌,并保持了与原始样品最相似的微生物群落结构。

在储存的样品中共检测到919种代谢物,其中633种处于正离子模式,286种处于负离子模式。按相对含量排在前十位的代谢物如图5A所示,其中脂质和类脂分子所占比例最大,其次是有机酸和衍生物。在贮藏结束时,CON、AG、CH和CE中的类脂和类脂分子的百分比分别为53%、36%、37%和27%。这表明脂肪代谢在贮藏过程中起主导作用,壳聚糖包被EGCG显著减缓了脂肪的降解。这与在每组中观察到的LOX活性相对应,并且可能与EGCG对LOX活性和腐败微生物的额外抑制有关。PCA结果(图5B)显示样本组之间的代谢差异显著,随着时间的推移,这种差异变得更加明显。15天后,CE组的代谢谱与最初的最相似样本,然后是CH、AG和CON组。这表明CE涂层有助于保持与初始样品更接近的代谢模式。分析了选定组的代谢差异,前20种上调和下调的代谢物显示在表S5-S10中,以及它们相应的VIP、P和FC值。为了便于更清楚地可视化,代谢差异用火山图表显示(图5C和图5C)。S3)和火柴棒图(图5D和图5D)。S4)。贮藏后,CON组IMP、尿苷、一磷酸鸟苷、S-腺苷同型半胱氨酸、肌苷、鸟苷、鸟苷和尿苷显著下调,黄苷和1,3-二甲基尿嘧啶显著上调,均与核苷酸代谢密切相关。此外,谷氨酸、组胺、身体碱、色胺和苯丙酮酸的表达显著上调,而D-(−)-谷氨酰胺、L-丝氨酸、氧化谷胱甘肽和L-甲状腺素的表达下调,均与氨基酸代谢密切相关。PC(10:0/10:0)、PC(7:0/13:1)、PC(9:0/13:1)、PC(8:0/13:1)和16-羟基十六酸显著下调,而硬脂酰乙醇酰胺、油酸、油酰乙醇胺和13,14-二氢前列腺素E1表达上调,与脂代谢密切相关。在碳水化合物代谢方面,D-红-4-磷酸、D-七氢呋喃7-磷酸和乳糖酸显著下调,而三羧酸(TCA)循环中的关键中间产物DL-苹果酸显著上调。与CON组相比,CE组显示与氨基酸代谢相关的代谢产物尸胺和组胺、脂代谢相关的代谢产物包括硬脂酰乙醇胺、11(Z)、14(Z)-二十碳二烯酸、11(E)-二十碳烯酸、油酰乙酰胺、油酰乙醇胺、二十二碳六烯酸、FAHFA(20:4/20:3)、二十二碳六烯酰乙醇胺、PC(14:0E/5:0)和反式石油硒酸,以及核苷酸代谢相关的代谢产物Xanthite Xanthosine、碳水化合物代谢相关的NADH和D-α羟基酒石酸(施等,2022)。这表明,CE涂层潜在地调节了关键的生化途径,导致氨基酸、脂肪、核苷酸和碳水化合物代谢的降低,从而有助于保存贮藏鲟鱼的风味品质和新鲜度。值得注意的是,与CH组相比,CE组第15天油酰乙胺、LPE 19:0、LPE 20:0、LPE 18:0、LPE 20:1、11(Z)、14(Z)-二十碳二烯酸、11(E)-二十碳烯酸、反式-石油硒酸以及PC(14,0e/5:0)和PC(5,0/13:1)的表达显著降低。这表明,EGCG的加入显著减缓了脂肪代谢,这可能是由于EGCG具有强大的抗氧化性和抑制脂氧合酶活性的能力。进行了偏最小二乘判别分析(PLS-DA),以更好地区分样本之间的代谢特征(图5E和S5A-S5E)。结果表明,不同组之间存在明显的差异,表明贮藏时间和包衣处理都显著影响了样品的代谢。此外,还评估了验证试验的R2和Q2值,以确保PLS-DA模型在识别关键标记代谢物方面的可靠性和准确性(图5F)。此外,还进行了KEGG途径浓缩分析,以进一步探索可能受到影响的代谢途径(图5G)。CON组和CE组之间的代谢比较显示,在几个途径上发生了显著的变化。值得注意的是,不饱和脂肪酸的生物合成是脂肪代谢的关键成分,显示出最显著的影响。这一途径,突出了它的大圆圈大小和深红色的地块,表明壳聚糖-EGCG涂层强烈抑制脂肪酸代谢。其他显著改变的代谢途径包括嘧啶代谢、乙醛代谢和二羧酸代谢,这表明壳聚糖-EGCG涂层不仅影响脂肪代谢,而且还影响核苷酸和碳水化合物的代谢。此外,赖氨酸降解和磷酸戊糖途径显示出较温和的变化,这表明涂层也可能影响氨基酸代谢和细胞氧化还原平衡。CH和CE基团间KEGG通路的丰富分析提出CE组中受影响最显著的途径是与脂代谢相关的途径,包括类固醇激素生物合成、不饱和脂肪酸生物合成和花生四烯酸代谢。这些途径对于脂类新陈代谢和生物活性脂类的产生是必不可少的。总而言之,这些发现表明,含有EGCG的壳聚糖涂层显著改变了多种代谢途径,特别是与脂质代谢相关的途径。这一改变可以有效地减少挥发性异味化合物的产生,从而提高鲟鱼在贮藏过程中的风味品质。
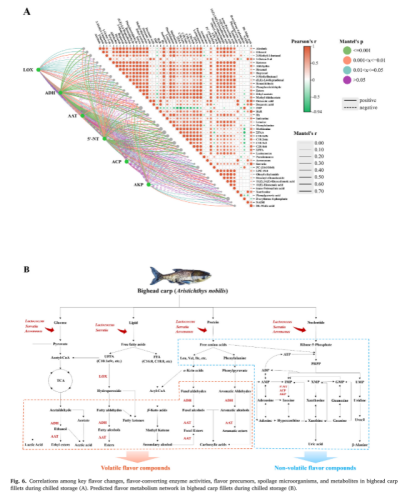
在图6A的右侧,主要风味化合物与微生物或代谢物之间的配对比较是使用颜色渐变显示的。正相关由红色圆圈表示,负相关由绿色圆圈表示,较强的相关性由较深的颜色或较大的圆圈表示。结果表明,关键的醇、醛、酮、酯和有机酸与FFA和FAA之间存在很强的正相关关系,这可能是因为FFA和FAA是这些挥发性化合物的潜在前体。这些挥发物与乳球菌和沙雷氏菌的存在之间也存在显著的正相关,这表明这两种微生物可能是储存过程中腐败气味的主要贡献者。3-甲基-1-丁醇和3-甲基丁醛与支链氨基酸亮氨酸和异亮氨酸以及沙雷氏菌与1-辛烯-3-醇之间均呈显著正相关。此外,亚油酸与(E,E)-2,4-庚二烯有很强的正相关关系,类似于以前的发现。苯甲醛和苯乙醛与苯丙氨酸呈强正相关,而蛋氨酸与硫代乙酸甲酯呈正相关。IMP与乳球菌、沙雷氏菌和气单胞菌呈负相关,而与HxR、HX和Xanthosine呈正相关,表明它们可能是IMP降解和HxR、Hx和Xanthosine产生的主要贡献者。除PC(10:0/10:0)外,其他主要脂代谢产物与脂肪族挥发性化合物、乳球菌和沙雷氏菌呈正相关。此外,NADH与气单胞菌呈正相关,而DL-苹果酸与乳球菌、沙雷氏菌和气单胞菌呈正相关,提示这些属可能直接影响碳水化合物的代谢。图6A的左侧接受了曼特尔测试,以评估酶、风味物质、腐败微生物和代谢物之间的相关性。连接线根据曼特尔p值进行颜色编码,其粗细代表相关性的强度。脂肪氧合酶D与脂肪醇和醛,特别是乙醇、己醛、庚醛和(E,E)2,4-庚二烯呈显著正相关(p≤0.05)。此外,抗酸脱氢酶与酒精,特别是乙醇和3甲基-1-丁醇之间存在高度显著的正相关关系(p≤0.001),并且与酯类之间存在一定程度的缔合关系。5‘-NT与IMP、HxR和Hx有较强的相关性,而AKP和ACP与这些代谢物的相关性相对较弱,表明5’-NT可能是对核苷酸代谢影响最大的酶。总而言之,鲤鱼样品中的风味变化与特定的酶、腐败微生物和风味前体密切相关。相关分析和KEGG数据库使之能够构建鲟鱼的风味代谢网络(图6B),其中脂肪、蛋白质、核苷酸和碳水化合物的代谢被确定为主要途径。在上一节观察到的代谢物调节的基础上,壳聚糖基涂层通过影响四个关键的代谢途径改善了鲟鱼样品的风味品质。值得注意的是,与纯壳聚糖涂层相比,添加EGCG的壳聚糖具有更明显的抑制脂肪代谢和减少脂肪族异味化合物产生的效果,进一步提高了风味品质。这种效果可能是由于EGCG具有强大的抗氧化性能,以及它抑制酶活性和抑制特定腐败微生物的增殖的能力。上述研究结果证实了壳聚糖没食子儿茶素没食子酸酯涂层可以通过抑制风味相关酶的活性,抑制腐败微生物的增殖,减缓蛋白质、脂类和核苷酸的降解,显著改善冷藏鲟鱼鱼片的整体风味品质。
总结
研究结果表明,添加EGCG的壳聚糖涂膜能有效地改善鲟鱼鱼片在4◦C贮藏过程中的感官品质,减少异味物质的产生。该涂层抑制挥发性醛、醇、酮、令人不快的FAAS、HX、HxR和关键酶(LOX、ADH、5‘-NT)的活性,以及腐败微生物(如乳球菌、沙雷氏菌、气单胞菌)的生长。代谢组学分析揭示了脂肪代谢途径的显著调节,相关分析突出了异味产生、风味前体、酶和腐败微生物之间的正向联系。这些效应归因于EGCG的抗氧化剂和抗真菌特性,为了解水产品的风味恶化机制提供了见解,并提出了改善水产品货架期和质量的潜在策略。
原文DOI:10.1016/j.foodchem.2025.143536
来源:公众号-食品指南针
原文链接:https://mp.weixin.qq.com/s/wIss1oy_3EHPeRDL-0Fgpw


